
Fosilni dokazi navodnih ljudi-majmuna—2. dio: ne-Homo hominidi
Cilj ovog pregleda je kritički analizirati fosilne dokaze za navodne ljude-majmune koji nisu klasificirani u rodu Homo. Svaki od ovih kandidata za hominide bit će ispitan otprilike ‘evolucijskim’ kronološkim redom, počevši od najranije. Morfologija kostura ovih navodnih ljudi-majmuna ukazuje da su se radi o izumrlim čovjekolikim majmunima koji nisu imali nikakve veze s evolucijom čovjeka.
Tounaigb/illustragb.html

Navodni ljudi-majmuni pripadaju kategoriji fosila poznatoj kao hominidi, skupini koja uključuje žive ljude, njihove pretke i sva druga stvorenjima slična majmunima za koja se vjeruje da su u bližem srodstvu s ljudima nego s čimpanzama. Neki evolucionisti ovu skupinu sada nazivaju homininima.1 Izraz hominid besmislen je s kreacionističkog gledišta, jer su hominidi bili ili ljudi ili čovjekoliki majmuni, a ne nešto između. Izraz se ovdje koristi samo za kategorizaciju ovih fosila i ne implicira vjerovanje da su takva stvorenja stvarno postojala. Prema evolucionistima, odlučujući trenutak za određivanje hominida je hipotetski razlaz između ljudi i čimpanzi. U prošlosti je kalibracijska točka ljudi i čimpanzi određena molekularnim satom na prije 5 milijuna godina (Ma), koristeći za kalibracijsku točku navodno odvajanje majmuna starog svijeta i čovjekolikih majmuna prije 30 Ma, na osnovu konvencionalnog radiometrijskog datiranja fosila.2 Evolucionisti sada smatraju da se odvajanje dogodilo prije oko 5 do 7 Ma. Radiometrijsko datiranje3 i molekularni satovi4 se temelje na nedokazanim i problematičnim pretpostavkama. Kad se evolucijska starost fosila navodi u ovom članku, to je u svrhu njihovog stavljanja u evolucijski kontekst i ni na koji način ne implicira da su ti datumi valjani.
Australopiteci se odnose na članove roda Australopithecus, i u ranijim vremenima ovo je bio jedini rod hominida osim Homo. Međutim, posljednjih godina mnogi su hominidi svrstani u nove taksone van Australopithecusa, poput rodova Ardipithecus, Sahelanthropus, Orrorin i Kenyanthropus. Sa točke gledanja stvaranja, treba podsjetiti da vrsta nije ekvivalent biblijskoj vrsti.5
Sahelanthropus tchadensis
Ako je publicitet koji okružuje najavu pronalaska fosila nešto po čemu se treba ravnati, tada je Sahelanthropus tchadensis favorit za titulu najranijeg hominida, budući da je pompa bila izvanredna čak i za fosila ljudi-majmuna, koji imaju reputaciju da čine mnogo ‘buke’ pri dolasku. Lubanja Sahelanthropus (TM 266–01–060–1, nadimka Toumai), pronađena u Chad-u, centralna Afrika, prikazana je na naslovnici izdanja od 11. srpnja 2002 časopisa Nature, uz naslov ‘Najraniji znani hominid’. Lubanju i druge fosile (fragmentne donje čeljusti) istraživači su datirali, koristeći pridruženu faunu, na između 6 i 7 milijuna godina, s procijenjenim kapacitetom lubanje između 320 do 380 cm3.6 Čudna stvar u vezi lubanje Toumai je to što straga izgleda poput čimpanze, ali sprijeda, jedan ekspert kaže da prolazi kao napredni australopitek.7 Prema Bernard-u Wood-u, hominid te starodti ‘ne bi trebao imati lice hominida manje od jedne trećine svoje geološke starosti’.7 Zato, ako evolucionisti prihvate Sahelanthropus kao najraniji ogranak hominida, tada to baca legitimnu sumnju na status predaka svih ostalih hominida ‘primitivnijih’ lica,7 i kao takav, bi poništio većinu geološki mlađih australopiteka.
Gotovo prije nego što se tinta osušila na razvikanim naslovima koji okružuju Toumai-ev dolazak, drugi evolucionisti, uključujući Brigitte Senut sa prirodoslovnog muzeja u Parizu, izrazili su sumnju u Toumai-ov status hominida. U jednom intervjuu iznijela je svoje stajalište kao sklono ‘mišljenju da se radi o lubanji ženke gorile’.8 Ovo jedva da potvrđuje raniju grandioznu tvrdnju harvardskog paleontologa Daniel-a Lieberman-a da je Toumai ‘najvažnije otkriće fosila u živom sjećanju’.9 Tim kojeg je vodio Wolpoff tvrdio je kako su Toumai-eve impresivane supraorbitalne kosti, koje su veće no u drugih australopiteka i izumrlih čovjekolikih majmuna, vjerojatno mehanički odgovor na naprezanje,10 a ne indikator facijalne sličnosti s rodom Homo. Ispitivanje ožiljaka koje su na stražnjoj strani lubanje ostavili mišići vrata dovelo ih je do zaključka da je Toumai bio četveronožac (tj. hodao je na četiri noge, a ne na dvije).11 Nakon svog ispitivanja Toumai-a, iznijeli su mišljenje ‘da je Sahelanthropus bio čovjekoliki majmun’.12 Još jedna navodna osobina hominida Toumaija su njegovi mali očnjaci u usporedbi s postojećim čovjekolikim majmunima.13 Iako se Sahelanthropus po nekim odlikama razlikuje od gorila, postoje i mnoge sličnosti, te kako kaže Matthew Murdock postoji velika mogućnost da su Sahelanthropus i živuće gorile srodne u smislu da su pripadnici iste biblijske vrste.14 Ako je to točno, tada maleni očnjaci mogu odražavati nekadašnju veću raznolikost u veličini očnjaka ove vrste čovjekolikog majmuna, gdje je genetska informacija za male očnjake vremenom izgubljena.
Nedavno su se pojavile tvrdnje da je jedan od kutnjaka u donjoj čeljusti zalijepljen na krivo mjesto, te da sjekutić prikupljen na tom mjestu nije prikazan u originalnom radu objavljenom u časopisu Nature.15 Autori tvrde da su fosili pripisani Sahelanthropusu ‘subjekt debate da li oni predstavljaju hominida ili čovjekolikog majmuna. Iz tog je razloga neophodno pružiti potpune i točne detalje vezane za taj fosil.’16 Kao što se često događa u paleoantropologiji kada je establišment doveden u pitanje, par koji je iznio tvrdnje pretrpio je odmazdu.17 Također je bilo predvidljivo da će prebacivanje Sahelanthropusa u čovjekolike majmune pratiti poznat obrazac ne primanja niti približne medijske pažnje kakva je bila ona u slučaju njegova prvotnog uzdizanja u hominide.
Orrorin tugenensis
Početkom 2001. Orrorin tugenensis, za koji se tvrdi da je živio prije oko 6 Ma, proglašen je kandidatom za najranijeg hominida (fosilne kosti su pronađene u Keniji, Tugen Hills), od strane tima kojeg su vodili Brigitte Senut i Martin Pickford.18 Fosili su se sastojali od 13 dijelova, uključujući slomljene bedrene kosti, nekoliko zuba i dijelove donje čeljusti.19 Stoga nije moguće znati kako je izgledala glava ovog stvorenja. Jedini razlog zbog kojeg Orrorin polaže pravo na status ranog hominida, kao i tvrdnje o dvonožnosti, jest da je pronađen u odgovarajućem evolucijskom vremenskom razdoblju. Evolucionisti smatraju bipedalizam važnim dokazom statusa hominida. Orrorin je navodno hodao na dvije noge zbog nakošene bedrene kosti koja nalikuje na onu čovjeka, temeljeno na dugom vratu bedrene kosti i žlijebu na stražnjoj strani tog vrata koji je prisutan u ljudi, ali ne u čimpanzi.20 Međutim, taj žlijeb, gdje mišić obturator externus pritišće o kost, također postoji i u stvorenja koja nisu dvonožna,21 te prema jednom ekspertu, skeniranja bedrenog vrata Orrorina pokazuju distribuciju kortikalnog koštanog tkiva poput onog u čimpanzi.20 Drugi argument je taj da je glava bedrene kosti Orrorina proporcionalno veća od one u Lucy, a evoluiranje takve bedrene kosti bi pomoglo raspodjeli sile uzrokovane dvonožnim kretanjem.22 Tim je kontroverzno predložio da je Orrorin zasebno razvio dvonožnost, neovisno o Lucy i drugih vrsta Australopithecusa, koje oni prebacuju u izumrli ogranak hominid porodice.22
Orrorin kontroverza nastavljena je u rujnu 2004. godine objavom rezultata računalne tomografije (CT) bedrene kosti, i tvrdnje o ne-majmunskoj, kasnije hominid, distribuciji kortikalnog koštanog tkiva u vratu bedrene kosti Orrorina, što upućuje na dvonožno kretanje.23 Kako je izvijestila Ann Gibbons, govoreći o ovim najnovijim nalazima:
‘Senut je pretpostavio da je Orrorinov hod sličniji ljudskom nego hodu australopiteka starih 2 do 4 milijuna godina. Ako je tako, australopiteci bi bili izbačeni iz izravne linije ka ljudima—dramatična revizija naše prapovijesti.’24
Paleontolog Tim White nije se složio s tom procjenom ‘hoda’ te napao najnovija objavljena otkrića rekavši da je ‘rezolucija CT-a toliko loša da nije bilo moguće biti siguran u uzorak debljine kosti’.24 White je također ‘nazvao Senut-ovo premještanje australopiteka “une position créationniste”, jer sugerira da je Orrorin-ova bedrena kost bila sasvim moderna prije 6 milijuna godina, a ne da se razvijala u fazama’.24 Čini se da čak i evolucionisti koji odlutaju izvan ortodoksne priče o evoluciji hominida riskiraju odmazdu.
Prema David-u Begun-u, University of Toronto, dokaz dvonožnosti Orrorina je dvosmislen.21 Ubrzo nakon prve objave fosila, Begun je komentirao da fragmenti fosila koji predstavljaju Orrorina ne mogu otkriti je li on ‘na liniji s ljudima, na liniji s čimpanzama, zajednički predak oboma ili samo izumrla bočna grana’.19
Ardipithecus kadabba
Nekoliko mjeseci nakon što je predstavljen Orrorin tugenensis, još je jedan hominid kandidat, iz Middle Awash-a u Etiopiji, nazvan Ardipithecus ramidus kadabba, datiran na 5.2 i 5.8 Ma, opisan u časopisu Nature.25 Taj je primjerak, 2004. godine, podignut iz statusa pod-vrste u zasebnu vrstu, Ardipithecus kadabba, na temelju pronalaska dodatnih fosiliziranih zubi.26 Kod kadabba-e (kao i kod Orrorin-a) fosilni su se nalazi sastojali od nešto post-kranijalnih kostiju, zubiju i fragmenata čeljusti, pa nije poznato kako je izgledala glava tih stvorenja.
Kadabba je interpretiran kao dvonožac na temelju karakteristika jedne kosti nožnog prsta (proksimalna falanga), odnosno dorzalne orijentacije površine proksimalnog zgloba.27 Argument je taj da je zglobna površina nožnog prsta nakošena prema gore slično kao u ljudi, dok je u čimpanzi nakošena prema dolje, te je tako navodno dokaz da je kadabba hodao poput ljudi.28 Međutim, kao što naglašava Begun,
‘ista se konfiguracija zglobova javlja kod definitivno ne-dvonožnog kasnog miocenskog hominida Sivapithecus, te duljina i zakrivljenost ove kosti vrlo nalikuje onima čimpanze ili bonoba’.29
Nevjerojatno, kost nožnog prsta datirana je kao nekoliko stotina tisuća godina mlađa od ostalih fosila, a pronađena je na lokalitetu udaljenom 16 km od ostalih, zbog čega čak i poznati lovac na hominide Donald Johanson sumnja u kategorizaciju kosti nožnog prsta s ostatkom fosila.30
Na osnovu nekoliko (jedanaest) fosilnih ostataka, koji predstavljaju najmanje pet različitih jedinki, s pet različitih lokacija,25 čini se nevjerojatnim da se svi mogu označiti kao pripadnici iste podvrste, a kasnije, na temelju pronalaska nekih dodatnih zuba,26 da se fosili mogu pripisati novoj vrsti. Prema Balter-u i Gibbons-u, ‘Timovi Orrorin-a i Ardipithecus-a tvrde da bi fosili onih drugih mogli predstavljati pretka čimpanzi ili drugih čovjekolikih majmuna, a ne jednog od naših ranih ljudskih predaka ili rođaka.’31 Možda su oba tima djelomično u pravu, jer postoji malo sumnje da su oba hominida bili obični čovjekoliki majmuni.
Ardipithecus ramidus
1994. godine Tim White i njegov tim objavili su pronalazak Australopithecus ramidus-a iz Aramis-a, Etiopia, koji je, u to vrijeme, datiran na 4.4 Ma, bio najraniji ‘hominid’ pronalazak od bilo kakvig značaja.32 Oko osam mjeseci kasnije, u ispravci, autori su reklasificirali fosile u potpuno nov rod, Ardipithecus, s novim imenom vrste corrigendum Ardipithecus ramidus.33 Izvorni članak je popraćen fotografijom, na naslovnici Nature-a od 22 rujna 1994, malog fragmenta čeljusti Ardipithecus ramidus-a, koji sadrži zub, s naslovom ‘Najraniji hominidi’. Stoga, netko bi očekivao značajan pronalazak, no umjesto toga fosil, koji se sastoji od postkranijalnih, zubnih i kranijalnih ostataka, prema riječima samog autora, ‘pokazuje mnoštvo osobina koje se obično povezuju s modernim čovjekolikim majmunima’.34 Autori zagovaraju status hominida uglavnom na temelju morfologije očnjaka koja više nalikuje onoj sjekutića i više anteriornog položaja foramena magnuma, jer vjeruju da ‘Stjecanje ovih osobina kod Aramisa može biti u korelaciji s dvonožnošću, iako to tek treba demonstrirati.’35 Čak i u evolucionističkoj zajednici, nisu svi uvjereni da je ramidus hominid. Prema Peter-u Andrews-u, pri London’s Natural History Museum, tanka caklina na zubima ramidus-a ‘je više u skladu s onime što bi očekivali od fosila čimpanze’, a osobine kosti ruke ‘sugeriraju četveronožno hodanje u stilu čimpanze’.36 Nedavno je više fosilnih ostataka iz Afara u Etiopiji (čeljusti, zubi, nožni prsti i kosti prstiju) dodijeljeno najmanje devet jedinki ramidus-a, a tvrdnje o dvonožnosti iznesene su na temelju dorzalne orijentacije proksimalne površine zgloba proksimalne falange stopala, osobine koja se također vidi u kadabba-e.37 Za opovrgavanje ove tvrdnje o dvonožnosti vidi kadabba ulomak iznad u tekstu.
Australopithecus anamensis
1995. godine, dentalni, kranijalni i postkranijalni uzorci sa dva odvojena lokaliteta u Keniji, datirani na oko 3.8 do 4.2 Ma, objavljeni su kao pripadnici nove vrste hominida, Australopithecus anamensis.38 Većina fosilnih ostataka nedvojbeno je potjecala od čovjekolikog majmuna, poput čeljusti nalik onoj čimpanzi, ali kontroverze su okruživale navodnu ljudsku prirodu tibije i humerusa.39 Humerus anamensisa nema duboku, ovalnu šupljinu, koja služi kao mehanizam zaključavanja humerusa i ulne, potonja je prisutna kod čimpanza, ali ne i kod ljudi, a tibia anamensisa je široka, kao i kod ljudi, zbog dodatnog spužvastog tkiva, koji djeluje kao amortizer tijekom dvonožne lokomocije.40 U vrijeme objavljivanja, paleontolog Peter Andrews iznio je sugestiju da bi tibija i nadlaktična kost, koje dolaze iz slojeva (gornji nivo) lokaliteta Kanapoi različitih od onih iz kojih dolaze ‘primitivne’ čeljusti i zubi (niža razina), mogle ‘ biti povezane s ljudima, a druge čovjekolikim majmunima’.41 Bilo je nekih fragmenata mandibule (KNM-KP 29287) pronađenih u slojevima gornje razine, no oni su bili iz druge (više) razine od tibije (KNM-KP 29285) i nadlaktične kosti (KNM-KP 271).42 U to vrijeme također je postojao problem od nekoliko stotina tisuća godina evolucijskog vremena između gornje i donje razine, ali u kasnijem radu dobiveni su datumi koji su suzili vremenski jaz.43 U ovom kasnijem članku prijavljeni su dodatni nalazi fosilnih ostataka iz anamensisa, ali nisu pruženi dodatni dokazi koji bi ih povezali s tibijom i humerusom. Također, analiza radijusa fosila iz 2000. (KNM-ER 20419) s druge anamensis lokacije, Allia Bay, ukazala je na ‘specijaliziranu morfologiju zapešća povezanu s hodanjem na zglobovima’.44 U radu čiji je koautor Meave Leakey, kaže se da je anamensis veoma sličan afarensisu, postkranijalno, s humerusom, tibijom i radijusom koji ‘gotovo točno odgovaraju po veličini i morfologiji zbirkama A. afarensisa iz Hadara’.45 Dakle, ako postkranijalni ostaci pripadaju anamensisu, tada u najboljem slučaju imate majmunoliko stvorenje koje se kreće na sličnim način kao majmunoliki afarensis. Budući da je provedena dublja analiza afarensisa, kojemu se pripisuje više postkranijskih fosila, čitatelja upućujemo na taj odjeljak.
Australopithecus afarensis

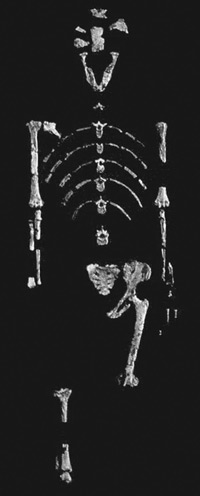
Otkriće slavnog Lucy-nog kostura u Etiopiji 1974. godine46 i kontroverza oko imenovanja Australopithecus afarensis 1978.47 dobro su poznati i nećemo ih ovdje prepričavati. Procijenjeni kranijalni kapacitet afarensisa, čiji je evolucijski vremenski raspon bio od oko 3.0 to 3.9 Ma, je između 375 i 540 cm3, s prosjekom od oko 470 cm3,48 iako je srednja veličina mozga drugdje navedena kao 438 cm3.49 Iznad vrata, lubanja (uključujući čeljusti) afarensisa opisana je kao majmunolika,50 a stručnjaci za evoluciju razlikuju njezinu lubanju od lubanje čimpanza, uglavnom zbog manjih očnjaka, većih zuba iza očnjaka afarensisa, ‘i utjecaja koji manji očnjaci imaju na lice A. afarensisa, uključujući smanjenu njušku i prisutnost fossa canina’.51 Detalji o prvoj lubanji afarensisa (AL 444–2) objavljeni su 1994. godine,52 i u to vrijeme bila je to najveća poznata lubanja australopiteka,53 s zapreminom procijenjenom na preko 540 cm3.50 Prethodno je rekonstrukcija lubanje afarensisa bila kompozitna, na osnovu fragmenata više jedinki,54 te je kao rezultat toga bilo optužbi da su Kimbel i White, koji su načinili kompozitnu lubanju, ‘na silu prilagodili vitko lice Homo-like vrste na koštani ovoj mozga neke robusne vrste’.55
Postkranijalno, afarensis je posjedovao majmunolike osobine poput koničnog prsnog koša i poput drugih australopiteka, bio snažne građe za svoj stas, te ‘te gotovo sigurno nisu bili, poput ljudi, prilagođeni uspravnom hodu i trčanju’.56 Ljudi imaju rebra u obliku bačve, a u usporedbi s čovjekolikim majmunima, lagani su za svoj stas. Analiza čašice lopatice (scapula) afarensisa pokazala je da je bila usmjerena više kranijalno (prema gore) u odnosu na ljude,57 čija je čašica okomita prema tlu, te da bi ta, kao u majmuna, orijentacija čašice afarensis prema gore bila ‘korisna da su ruke držane iznad glave velik dio vremena, kao što je slučaj tijekom penjanja na stabla i vješanja’.58 Što se tiče proporcija udova, procijenjeni humerofemoralni indeks (omjer duljine humerusa i bedrene kosti) kod afarensisa je manji nego kod postojećih čovjekolikih majmuna, iako je još uvijek značajno veći od onog kod ljudi.59 Ljudi imaju relativno kratke ruke i vrlo duge noge, a prema Tattersall-u i Schwartz-u, ‘činjenica je da su noge afarensisabile prilično kratke’.60 Procijenjeni brahijalni indeks (omjer radijusa i duljine nadlaktične kosti) kod afarensisa je veći nego kod ljudi i gorila, ali manji nego kod drugih postojećih čovjekolikih majmuna, no ovaj procijenjeni indeks je vrlo nepouzdan zbog poteškoća u izradi pouzdane procjene dužine radijusa Lucy (AL 288–1), a velikodušna procjena dužine radijusa dovela bi brahijalni indeks unutar raspona čimpanze.61 Međutim, Tattersall i Schwartz izjavljuju
‘čak i ako su relativne proporcije ruku sličnije ljudskim nego majmunskim (donji ud je mnogo duži u usporedbi s gornjom u čimpanzi, a posebno u pravih arborealista—orangutana i gibona)—ruke afarensisa su još bile prilično duge’.60
Kosti prstiju ruku afarensisa, osobito proksimalne falange, zakrivljene su, slično onima u čimpanza i ‘ukazuju na prilagodbu za aktivnosti suspenzije i penjanja koje zahtijevaju snažne sposobnosti hvatanja’.62 Nožni prsti afarensisa također su zakrivljeni,63 kao kod čovjekolikih majmuna, i sugeriraju sposobnost penjanja po drveću. Prema Sternu i Susmanu, ostaci stopala i gležnja afarensisa ‘otkrivaju nam životinju koja se penjala, ali i bila dvonožna’.64 Dalje navode da ‘Nema dokaza da bilo koji postojeći primat ima dugačke, zakrivljene, mišićave šake i stopala za bilo koju drugu svrhu osim za ispunjavanje zahtjeva punog ili povremenog života na stablima.’64
Analiza fosila ruku afarensis od strane Sterna i Susmana 1983. godine ukazala je na mnoge značajke koje su zajedničke čimpanzama, ali su istaknuli da ispitivanje metakarpala nije dalo dokaze da afarensisje hodao na zglobovima.65 Dokazi da je afarensis bio hodač na zglobovima kasnije će se, međutim, pojaviti. Morfologija zapešća kod postojećih afričkih čovjekolikih majmuna koji hodaju s zglobovima prstiju (čimpanze i gorile) ima specijalizirani mehanizam za zaključavanje koji ograničava ‘istezanje zgloba tijekom oslanjanja pri hodanju na zglobovima prstiju’.44 Godine 2000. Richmond i Strait izvijestili su da je distalna radijalna morfologija (uključujući skafoidni usjek) kod anamensisa i afarensisa slična onoj kod zglobova afričkih čovjekolikih majmuna, dok je radijus pripisan robustusu, a posebno africanusu, bio sličniji čovjeku.66 Zapanjujuće, autori su sugerirali da su značajke hodanja na zglobova kod anamensisa i afarensisa nefunkcionalno nasljeđe od ranijeg pretka.67 Međutim, nefunkcionalna zadržavanja (evolucijska prtljaga) nisu nešto za što se očekuje da će dugo trajati, prema evolucionistu Jeremyju Cherfasu:
‘Sve o njezinu kosturu, od vrhova prstiju ruku do nožnih prstiju, sugerira da su Lucy i njezine sestre zadržale nekoliko osobina koje bi bile vrlo prikladne za penjanje po drveću. Neke od tih istih prilagodbi za penjanje po drveću još uvijek se mogu uočiti, iako znatno smanjene, kod mnogo kasnijih hominida kao što su 2 milijuna godina stari primjerci Homo habilisa iz klanca Olduvai. Moglo bi se tvrditi da su Lucyne prilagodbe za život na stablu samo ostatak iz njezine prošlosti, ali životinje često ne zadržavaju osobine koje ne koriste, a pronaći te iste značajke u primjercima 2 milijuna godina kasnije čini to vrlo malo vjerojatnim.’68
Još jedna zanimljivost iz Richmond i Strait studije bila je da je morfologija zapešća kod africanusa ‘modernija’ nego kod afarensisa. Međutim, druge su studije pokazale da su proporcije stopala, potkoljenice i udova kod africanusa više majmunolike nego kod afarensisa, iako je lubanja u afarensisa sličnija čimpanzi nego u africanusa.69 Netko bi se mogao zapitati kakav je navodni evolucijski odnos između africanusa i afarensisa su u ovoj kompliciranoj slici, budući da je afarensis predložen kao predak africanusa. Prema Collardu i Aiellu:
‘Više nije slučaj da lubanja ukazuje na jedan skup filogenetskih odnosa, a postkranijalni kostur —sve osim lubanje—na drugi. Umjesto toga, različiti dijelovi postkraniuma možda ne podržavaju istu filogenetsku hipotezu.’69
U pogledu na Lucy, prema profesoru arheologije Clive Gamble-u:
‘Anatomija pokazuje potpuno dvonožni hod, ali s nekim razlikama u odnosu na moderne ljude. Doista, oni su toliko distinktivni da je Charles Oxnard u svojoj iscrpnoj studiji udova zaključio da način kretanja nije ni majmunski ni ljudski. Niti je to bila prijelazna faza, već uistinu jedinstvena.’70
Središnji argument za to da je afarensis hominid je da je stvorenje imalo uspravno držanje i da je bilo sposobno za dvonožno kretanje. Argument da je afarensis bio dvonožac temelji se na skeletnim rekonstrukcijama zdjelice i kostiju donjih udova. Kako je pronađeno do 40% Lucynog postkranijalnog kostura, velik dio morfoloških i biomehaničkih analiza temelji se na tim kostima, iako ne isključivo. Jedan problem s rekonstrukcijama Lucy-inog kostura (datiranog oko 3.2 milijuna godina) je subjektivna priroda rada, budući da unaprijed određena uvjerenja o posturi afarensisa mogu utjecati na rekonstrukciju kostura. Kako inače sljedeća izjava Mauricea Abitbola, koji je proučavao Lucynu zdjelicu, može imati smisla?
‘Prevladavajuća gledišta o Lucynom držanju gotovo je nemoguće pomiriti. Kad se pogleda rekonstrukcija koju su predložili Lovejoy (1998) i Weaver et al. (1985.), stječe se dojam da bi njezina rekonstrukcija imala bila tijelo savršeno modernog ljudskog dvonošca (Slika 1a). Ali kada se pogleda preliminarna rekonstrukcija nedavno prikazana u Smithsonianu, stječe se dojam čimpanze koja nespretno pokušava stajati na stražnjim udovima i sprema se pasti na prednje udove (Lewin, 1988.). U potonjem, implikacija je “primitivan” oblik dvonožnosti kod hadarskih hominida. Da bi se riješile takve razlike, potrebno je više anatomskih (fosilnih) dokaza. Trenutno dostupni podaci otvoreni su za vrlo različita tumačenja.’71
Nevjerojatno, Lucyno držanje može se protumačiti kao bilo što, od modernog čovjeka do čimpanze. Stoga kreacionisti imaju puno pravo biti krajnje skeptični prema tvrdnjama da su Lucy i drugi australopiteci bili na stupnju evolucije između čovjeka i majmuna, kada se glavni dokaz za tu tvrdnju temelji na tako spekulativnoj interpretaciji podataka. Iz svoje studije, Abitbol je zaključio da je ‘Lucyno uspravno držanje drugačije od onoga koje se može vidjeti kod modernih ljudi i da je još uvijek misterij’.72
Još jedan problem sa studijama koje istražuju sposobnost kretanja fosila koji pripadaju stvorenjima kao što je afarensis je taj što se u raspravama o mišićnim funkcijama ‘pretpostavlja a priori da su mišići imali obrazac sličan onom u ljudi’.73 U biomehaničkoj analizi Lucyne zdjelice i kostiju donjih udova, posebno rekonstrukcije glutealne muskulature, Christine Berge je izvijestila da bi ‘majmunolika glutealna organizacija nudila bolju sposobnost od ljudske organizacije’.73 Iz studije, Berge zaključuje
‘da se dvonožnost Australopitekusa mora razlikovati od one Homo. Ne samo da je Australopithecus imao manju sposobnost održavanja ekstenzije kuka i koljena tijekom hodanja, nego je vjerojatno i drugačije pomicao zdjelicu i donje udove. Čini se da se hod australopiteka značajno razlikovao od ljudskog, te je uključivao neku vrstu geganja, s velikim rotacijskim pokretima zdjelice i ramena oko vertikale (Berge, 1991.a, b). Takvo hodanje vjerojatno je zahtijevalo veći energetski trošak od ljudskog bipedalizma.’74
Iz svoje detaljne analize postkranijalnog kostura, Stern i Susman ‘otkrili su značajan broj dokaza koji pokazuju da su aktivnosti na drvetu bile toliko važne A. afarensisu da su morfološke prilagodbe koje omogućuju vješto kretanje po drveću održane’.75 Što se tiče afarensisa, također su zaključili ‘da je priroda zemaljske dvonožnosti, kada se prakticirala, bila drugačija od one modernih ljudi’.75 Također, CT skeniranja koštanog labirinta unutarnjeg uha (polukružni kanali ispunjeni tekućinom) koji je dio vestibularnog aparata odgovornog za ravnotežu, a koja je načinila grupa koju predvodi anatom Fred Spoor sa Sveučilišta u Liverpoolu,76 pokazala su da su dimenzije polukružnog kanala u lubanji koje se pripisuju drugim australopitecima (africanusu i robustusu) slične dimenzijama postojećih velikih majmuna.77 Iako skeniranje afarensisa nije provedeno, vjerojatno je da bi labirinti afarensisa nalikovali istom uzorku čimpanze/čovjekolikog majmuna kao kod australopiteka.78 Dakle, dokazi pokazuju da je vrlo malo vjerojatno da je uobičajeni način hodanja afarensisa bio dvonožan. Također nije jasno je li ograničeni dvonožni pokret koji pokazuju neki australopiteci, kao što je afarensis, bio posturalan i povezan s hranjenjem, nasuprot kretanju.79 Treba imati na umu da je za postojeće bonobe (pigmejske čimpanze) uočeno da hodaju dvonožno oko 10% vremena, tako da se ograničeni neljudska dvonožnost javlja čak i kod postojećih majmuna.79
Dakle, što točno imamo u afarensisu? Odgovor je stvorenje s mozgom veličine majmuna, majmunolikom lubanjom, s tijelom oblikom i veličinom sličnim majmunu, te stvorenje koje je bilo specijalizirano za penjanje po drveću i hodanje na zglobovima, slično majmunima. Drugim riječima, imate čovjekolikog majmuna. Međutim, ako je afarensis također imao ograničenu sposobnost neljudskog dvonožnog kretanja, koja je možda bila učinkovitija od one postojećih bonoba, je li to dokaz da je stvorenje bilo čovjek majmun ili samo odražava raznolikosti strukture tijela čovjekolikog majmuna/australopiteka koje je stvorio Bog?
Da odgovor leži u potonjem pokazuje analiza ostataka kostura čovjekolikog majmuna iz Italije nazvanog Oreopithecus bambolii, datiranog od 7 do 9 Ma, kojeg izgleda nitko ne želi označiti kao hominida, budući da su fosilni ostaci na pogrešnoj lokaciji i izvan ‘okvira’ hominidskog vremena, ipako bambolii ima anatomiju koja podržava ograničenu dvonožnu lokomociju.80 Prema autorima studije, dijelovi zdjelice bambolii-a podsjećaju na afarensis-a, a njegova bedrena kost pokazuje ‘naglašeni dijafizni kut u kombinaciji s kondilima podjednake veličine, sličan Australopithecusu i Homo i funkcionalno povezan s dvonožnim aktivnostima’.81 Prema Henry-ju Gee-u, ‘smatra se da je ovo stvorenje samostalno postalo dvonožno i da je samo u dalekom srodstvu s hominidima’.82 Dovoljno je teško povjerovati ili zamisliti da su čovjekoliki majmuni razvili oblik dvonožnog kretanja; da se to moralo neovisno dogoditi više puta, kako bi se ‘spasila’ evolucijska teorija otkriva evoluciju kao zbirku priča koje se mogu prilagoditi gotovo svakom scenariju, bez obzira koliko apsurdan ili malo vjerojatan bio. To da je ograničeni neljudski oblik dvonožnog kretanja postojao kod čovjekolikih majmuna, a koji nije povezan s bilo kojim hipotetskim hominidom i javlja se ranije od njih, ukazuje na to da posjedovanje dvonožnih osobina nije pokazatelj statusa hominida.
Kenyanthropus platyops
U ožujku 2001. još jedan ‘hominid’ pojavio se na naslovnici časopisa Nature, s naslovom ‘Ljudska se obitelj širi’. Članak je opisivao novo stvorenje, sa jezera Turkana, Kenija, zvano Kenyanthropus platyops.83 Datiranje na 3.5 Ma je dovelo platyops u izravnu konkurenciju s afarensis-om kao hominidom koji je navodno iznjedrio rod Homo.84 Najveće uzbuđenje bilo je uslijed jako iskrivljene lubanje. Procijenjeno je da je njegov kapacitet lubanje majmunskih proporcija, ali uglavnom zbog njegovog ravnog lica, neuobičajenog za navodnog hominida u toj fazi evolucije, te je primjerak pripisan novom rodu Kenyanthropus. Međutim, postoje jake indikacije da je ravno lice rezultat ozbiljne distorzije šireće matrice, što je artefakt procesa fosilizacije,85 te je tako platyops bez sumnje samo još jedan čovjekoliki majmun.
Australopitekus bahrelghazali
Godine 1995. mandibula ovog stvorenja, pronađena u Čadu, opisana je kao najsličnija savremenoj vrsti afarensis.86 Uzorak iz Čada, čija se starost procjenjuje na između 3 i 3.5 Ma, 1996. godine pripisan je novoj vrsti Australopithecus bahrelghazali. Procijenjeno je da se razlikuje od afarensis-a prema određenim morfološkim značajkama mandibule, korijena pretkutnjaka i debljine cakline pretkutnjaka. Međutim, tako oskudno otkriće teško opravdava stvaranje nove vrste. Nema razloga promatrati bahrelghazali kao bilo što osim čovjekolikog majmuna, vjerojatno iste vrste kao što je i afarensis.
Australopithecus africanus

Prosječna veličina mozga sedam primjeraka Australopithecus africanusaje 451 cm3, s rasponom od 425 cm3 do 515 cm3.87 Evolucionisti vjeruju da je africanus živio u Južnoj Africi od otprilike 2.5 do 3 Ma, iako je možda postojao do 2. Ma.88 Kranijalno, glavna razlika u usporedbi s afarensisom je u licu, gdje je lice africanusa šire licem i s manje izbočenom čeljusti.89 Također, u usporedbi s afarensis-om, africanus-ovadonja čeljust je robusnija, zubi iza očnjaka su povećani, a prednji zubi smanjeni.89
Australopithecus afarensis-a mnogi smatraju pretkom africanus-a, ali usporedbe proporcija udova dviju vrsta ukazuju na manje majmunoliku veličinu zglobova prednjih i stražnjih udova i duljinu udova u navodno starijem i kraniodentalno ‘primitivnijem’ afarensis-u.90 Prema autorima ovih studija, ‘To implicira da su se, u lozi hominida, proporcije udova mijenjale naprijed-natrag ili da je naš sadašnji pogled na odnose s hominidima previše pojednostavljen.’91 Umjesto toga, zbrka u vezi s hominidnim odnosima postoji jer nikada nije postojao evolucijski odnos između hominida. Prema Kleinu i Edgaru, africanus je imao vrlo majmunolik gornji dio tijela, s dugim, snažnim rukama pogodnim za penjanje po drvetu, ali je donji dio tijela bio ‘oblikovan za uobičajeno dvonožno kretanje po tlu’.92 Međutim, kao što je gore spomenuto, CT skeniranje koštanog labirinta unutarnjeg uha pokazalo je da su dimenzije polukružnog kanala u lubanji koje se pripisuju africanus-u slične onima u postojećih čovjekolikih majmuna.77 Stoga, ako je africanus na neki način hodao dvonožno, malo je vjerojatno da je to bio uobičajen način njegova kretanja.
Ranija analiza postkranijalne anatomije afarensisa i africanusa dovela je McHenryja do zaključka
‘da je (1) postkranijalni kostur ranog Australopitekabio jedinstveno različit od svih živućih majmuna i ljudi, (2) postkranijalni kostur ranog A. africanus-a je vrlo sličan A. afarensis-u unatoč stvarnim razlikama u njihovim lubanjama, zubima, rasporedu i starosti, i (3) postoji jasan stupanj lokomotorne prilagodbe kod Australopithecus-a koji se razlikuje od bilo kojeg živog oblika.’93
Dakle, ako su dvije vrste dijelile ‘jedinstvenu konfiguraciju koja implicira da su lokomotorne i posturalne prilagodbe bile vrlo slične i različite od bilo kojeg živog oblika’,94 tada je lokomotorno ponašanje africanus-abilo slično afarensis-u. Kao što je ranije naznačeno, afarensis je bio čovjekoliki majmun koji je hodao na zglobovima i bio izgrađen za arborealni način života, iako je možda imao ograničenu sposobnost dvonožnog hodanja na način koji nije sličan čovjeku, kao što su to činili majmuni koji nisu povezani ni s navodnom evolucijom hominida, kao što je Oreopithecus bambolii. Kao i kod svih ostalih vrsta australopiteka, africanus je imao mozak veličine majmuna, imao je majmunoliku lubanju i bio je sličan čovjekolikim majmunima po obliku i veličini tijela. Stoga je najvjerojatnije objašnjenje da je africanus bio vrsta izumrlih majmuna, a čak i među evolucionistima bilo je sugestija da se africanus i afarensis ‘trebaju smatrati podvrstama jedne vrste’.95
Australopithecus garhi
Godine 1999. za ostatke Australopithecus garhi, navodno 2.5 milijuna godina starog hominida iz Hata u etiopskom srednjem Awashu, rečeno je da se razlikuju od afarensisa ‘svojom apsolutno većom postkaninom denticijom i morfologijom gornje trećine pretkutnjaka sa smanjenom projekcijom meziobukalne linije cakline i manjom okluzalnom asimetrijom’.96 Međutim, gledajući vrlo nepotpune ostatke lubanje stvorenja, s kapacitetom lubanje procijenjenim na 450 cm3, teško je ne zapitati se jesu li razlike u nekoliko obilježja zuba prenaglašene, budući se ovaj primjerak sigurno mogao pripisati vrsti afarensis. Možda je kasniji evolucijski datum bio faktor. Bez obzira na afinitet prema afarensis-u, majmunolika priroda njegove lubanje čini da je garhi ništa više od čovjekolikog majmuna
Zanimljiviji su bili postkranijalni fosili (BOU-VP-12/1) pronađeni na tom lokalitetu, koje istraživači nisu dodijelili garhi-u, a imaju humano-femoralni indeks.97 Međutim, Bouri kostur (BOU-VP-12/1) također je imao procijenjeni brahijalni indeks veći od svih postojećih majmuna, osim orangutana (Pongo pygmaeus).98 Ova kombinacija duge podlaktice poput čovjekolikog majmuna i proporcija humerofemoralnih udova poput ljudskih čini zbunjujuć evolucijski odnos predak–potomak, jer bi to ‘sugeriralo obrat u brahijalnim proporcijama kod Bouri kostura (više majmunolikom) od stanja A. afarensis-a, nakon čega slijedi drugi obrat u H. ergaster-u (više nalik čovjeku)’.99
Robusni australopiteci
Robusni australopiteci konvencionalno se sastoje od vrste Australopithecus robustus (1.5 do 1.8 Ma) iz Južne Afrike i dvije istočnoafričke vrste Australopithecus boisei (1.3 do 2.3 Ma) i Australopithecus aethiopicus (2.3 do 2.7 Ma), s kombiniranim evolucijskim rasponom starosti za tri vrste od 1.3 do 2.7 Ma.100 Neki ih paleoantropolozi svrstavaju u zaseban rod, Paranthropus. Evolucionisti nijednu od robusnih vrsta australopiteka ne smatraju precima pravih ljudi, već sporednim ograncima koji su završili u slijepoj ulici,101 ‘jer su njihovi zubi i lubanje bili jako specijalizirani i zato što su koegzistirali s vjerojatnijim precima nakon 2.5 milijuna godina’.102 Zbog ‘njihovih ogromnih zuba za žvakanje i grube lubanje’ su nazvani robusnim australopitecima, iako su imali mali mozak i tijelo.103 Iz lubanja ovih stvorenja očito je da na njima nije bilo ničeg ljudskog.
Australopithecus aethiopicus
Poznata ‘Crna lubanja’ (KNM-WT 17000), s kapacitetom od oko 410 cm3, jedan je od tri ili četiri fosilna primjerka koji čine vrstu aethiopicus104 koju neki evolucionisti smatraju vjerojatnom vezom između afarensis-a i druge dvije robusne vrste, boisei-a i robustus-a105 Međutim, Crna lubanja ‘je ekstreman A. robustus/boisei tip s masivnim sagitalnom grebenom, a dva pronađena zuba su četiri ili pet puta veći od ljudskih kutnjaka’,106 pa iako je aethiopicus navodno starija vrsta od robustusa/boisei, njegova hiper-robustnost ga diskvalificira kao prijelazni oblik. Morfologija crne lubanje ima mnogo sličnosti s onom mladog mužjaka gorile,107 pa je moguće da gorile i aethiopicus pripadaju istoj biblijskoj vrsti. Trenutačno nema postkranijskih fosila koji se pripisuju aethiopicus-u, dok su druga dva (moguće tri) primjerka pripisana aethiopicusu mandibule.108
Australopithecus boisei
Prosječna veličina mozga Australopithecus boisei, na temelju šest primjeraka, iznosi oko 452 cm3, s rasponom od 390 cm3 do 500 cm3,109 iako druge procjene boiseija daju srednju vrijednost od 521 cm3.49 Kranijalno, boisei se opisuje kao ‘jedini hominin koji kombinira masivno, široko, ravno lice s neurokranijem skromne veličine’.110 Nema postkranijalnih ostataka koji bi se definitivno mogli pripisati boiseiju, ali iz Koobi Fora djelomični kostur i pojedinačne postkranijalne kosti neuvjerljivo su povezani s vrstom.110 Prema Woodu i Richmondu, ‘djelomični kostur karakteriziraju proporcije udova koje nalikuju na A. afarensis (Grausz et al. 1988), i manje su majmunoliki od onih A. africanus-a’.110 Dakle, ako je djelomični kostur (KNM-ER 1500) pripadao boisei-u, onda je možda imao slično lokomotorno ponašanje kao afarensis, i ‘kao A. afarensis, relativno dugi prednji udovi sugeriraju da lokomotorno ponašanje P. boisei sadrži arborealnu komponentu’.111
Australopithecus robustus

Veličina mozga Australopithecus robustus-a, koja se trenutačno može procijeniti za samo jedan primjerak (SK 1585), iznosila je 530 cm3, ali je novija procjena to spustila na 476 cm3.109 Kranijalno, robustus je bio sličan boiseiju, ali lice mu nije bilo tako veliko i široko.110 Dakle, ako je robustus imao ‘robusnu’ lubanju, onda je lubanja boiseijabila hiper robusna. U usporedbi s africanus-om, zubi za žvakanje i lice robustus-a su bili veći, iako su mu zubi očnjaci i sjekutići bili manji,112 a prema Kleinu i Edgaru, obje vrste
‘su imale vrlo majmunolike gornje dijelove tijela s dugim, snažnim rukama koje bi ih činile okretnim penjačima po drveću. Razlikovali su se od čovjekolikih majmuna prvenstveno po donjim dijelovima tijela, koji su bili oblikovani za uobičajeno dvonožno kretanje po tlu, te po zubima’.92
Prema Woodu i Richmondu, morfologija zdjelice i kuka za koju se vjeruje da je povezana s robustus-om nalikuje onoj kod afarensis-a i africanus-a (‘ gracilni’ australopitek), te sugerira da je ‘hod P. robustus-a vjerojatno nalikovao ‘gracilnom’ australopeku’.112 Kao što je ranije navedeno, hod afarensisa nije bio ljudski. Također, kao što je ranije spomenuto, CT skeniranje koštanog labirinta unutarnjeg uha pokazalo je da su dimenzije polukružnog kanala u lubanji koje se pripisuju robustususlične onima postojećih velikih čovjekolikih majmuna.77
Zaključak
Morfološke značajke fosila hominida pripisanih taksonima izvan roda Homo indiciraju da su bili obični izumrli čovjekoliki majmuni. Vrsta nije ekvivalentna biblijskoj vrsti, a nekoliko ne-Homo hominida može samo predstavljati varijaciju unutar jedne ili više vrsta čovjekolikih majmuna, baš kao što se vjeruje da fosilne vrste uključene u Homo, isključujući nevažeći takson Homo habilis, predstavljaju varijaciju unutar jedne ljudske vrste. Vrlo je moguće da su postojeći majmuni preživjeli ostaci ovih vrsta hominidnih vrsta majmuna, koji su izgubili velik dio svoje genetske raznolikosti. Općenito, tamo gdje je dostupan fosilni materijal, ta su hominidna stvorenja imala mozak veličine čovjekolikog majmuna, lubanje koje su bile poput čovjekolikog majmuna i tijela slična oblikom i stasom čovjekolikim majmunima. Morfologija stvorenja kao što je Australopithecus afarensis indicira da su bili specijalizirani za penjanje po drveću, kao i hodanje na zglobovima, kao i čovjekoliki majmuni. Mnogo se govorilo o karakteristikama kostura, što ukazuje da su neki australopiteci, uključujući afarensisa, mogli također imati ograničenu sposobnost neljudskog dvonožnog kretanja. Međutim, slična ograničena dvonožna sposobnost također je postojala kod čovjekolikih majmuna koji se ne smatraju hominidima, kao što je Oreopithecus bambolii, datirana na vrijeme prije pretpostavljenog razdvajanja čovjeka i čimpanze. Stoga, kako se osobina može koristiti kao argument za jedinstvenost svih hominida i njihov evolucijski odnos, kada osobina nije jedinstvena tim navodnim hominidima? To je kao da kažete da su se kitovi morali razviti od stvorenja poput nilskih konja jer oboje mogu plivati (ili oni vjeruju i u to?). Pretpostavljam da je to razlog zašto evolucionisti toliko vole riječ paralelno, kao u paralelnoj evoluciji. Nazovite to kako želite, ali je li to znanost?
Reference i bilješke
- Harrub, B. and Thompson, B., The Truth About Human Origins, Apologetics Press Inc., Montgomery, AL, pp. 8–10, 2003. Natrag na tekst.
- Gribbin, J. and Cherfas, J., The First Chimpanzee: In Search of Human Origins, Penguin Books, London, pp. 116–117, 2001. Natrag na tekst.
- Sarfati, J., Refuting Compromise, Master Books, Green Forest, AR, pp. 376–388, 2004; Lubenow, M.L., Bones of Contention: A Creationist Assessment of Human Fossils, Revised and Updated, Baker Books, Grand Rapids, MI, pp. 273–292, 2004. Natrag na tekst.
- Harrub and Thompson, ref. 1, pp. 99–134; Lubenow, ref. 3, pp. 222–235. Natrag na tekst.
- Lubenow, ref. 3, p. 187. Natrag na tekst.
- Brunet, M. et al., A new hominid from the upper Miocene of Chad, Central Africa, Nature 418:145–151, 2002. Natrag na tekst.
- Wood, B., Hominid revelations from Chad, Nature 418:134, 2002. Natrag na tekst.
- BBC News, Skull find sparks controversy, news.bbc.co.uk/hi/english/sci/tech/newsid_2125000/2125244.stm, 13 July 2002. Natrag na tekst.
- Montgomery, B., Man’s ancestors 7m years old, The Australian, 11 July 2002, p. 1. Natrag na tekst.
- Wolpoff, M.H., Senut, B., Pickford, M. and Hawks, J., Sahelanthropus or ‘Sahelpithecus’? Nature 419:581–582, 2002. Natrag na tekst.
- Ancient skull more ape than human? www.cnn.com/2002/TECH/science/10/09/ancient.skull/index.html, 10 October 2002. Natrag na tekst.
- Wolpoff et al., ref. 10, p. 582. Natrag na tekst.
- Brunet et al., ref. 6, p. 151. Natrag na tekst.
- Murdock, M., Sahelanthropus tchadensis—the ambiguous ape, Journal of Creation 18(3):114, 2004. Natrag na tekst.
- Beauvilain, A. and Guellec, Y.L., Further details concerning fossils attributed to Sahelanthropus tchadensis (Toumai), South African J. Science 100:142–144, 2004. Natrag na tekst.
- Beauvilain and Guellec, ref. 15, p. 142. Natrag na tekst.
- Dalton, R., Brickbats for fossil hunter who claims skull has false tooth, Nature 430:956, 2004. Natrag na tekst.
- Aiello, L.C. and Collard, M., Our newest oldest ancestor? Nature 410:526–527, 2001. Natrag na tekst.
- Balter, M., Scientists spar over claims of earliest ancestor, Science 291:1460, 2001. Natrag na tekst.
- Wong, K., An ancestor to call our own, Scientific American Special 13(2):9, 2003. Natrag na tekst.
- Begun, D.R., The earliest hominins—is less more? Science 303:1479, 2004. Natrag na tekst.
- Shipman, P., Hunting the first hominid, American Scientist 90(1), 2002; www.americanscientist.org/template/AssetDetail/assetid/18641, 13 August 2004. Natrag na tekst.
- Galik, K., Senut, B., Pickford, M., Gommery, D., Treil, J., Kuperavage, A.J. and Eckhardt, R.B., External and internal morphology of the BAR 1002′00 Orrorin tugenensis femur, Science 305:1450–1453, 2004. Natrag na tekst.
- Gibbons, A., Oldest human femur wades into controversy, Science 305:1885, 2004. Natrag na tekst.
- Halle-Selassie, Y., Late Miocene hominids from the Middle Awash, Ethiopia, Nature 412:178–181, 2001. Natrag na tekst.
- Halle-Selassie, Y., Suwa, G. and White, T.D., Late Miocene teeth from Middle Awash, Ethiopia, and early hominid dental evolution, Science 303:1503–1505, 2004. Natrag na tekst.
- Halle-Selassie, ref. 25, p. 180. Natrag na tekst.
- Wong, ref. 20, pp. 8–9. Natrag na tekst.
- Begun, ref. 21, pp. 1478–1480. Natrag na tekst.
- Lemonick, M.D. and Dorfman, A., One giant step for mankind, Time (South Pacific) pp. 58–59, 23 July 2001. Natrag na tekst.
- Balter, M. and Gibbons, A., Another emissary from the dawn of humanity, Science 293:189, 2001. Natrag na tekst.
- White, T.D., Suwa, G. and Asfaw, B., Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia, Nature 371:306–312, 1994. Natrag na tekst.
- White et al., ref. 32, p. 88. Natrag na tekst.
- White et al., ref. 32, p. 311. Natrag na tekst.
- White et al., ref. 32, p. 312. Natrag na tekst.
- Fischman, J., Putting our oldest ancestors in their proper place, Science 265:2011, 1994. Natrag na tekst.
- Semaw, S. et al., Early Pliocene hominids from Gona, Ethiopia, Nature 433:304, 2005. Natrag na tekst.
- Leakey, M.G., Feibel, C.S., McDougall, I. and Walker, A., New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya, Nature 376:565–571, 1995. Natrag na tekst.
- Leakey, M. and Walker, A., Early hominid fossils from Africa, Scientific American 276(6):63–64, 1997. Natrag na tekst.
- Leakey and Walker, ref. 39, p. 63. Natrag na tekst.
- Andrews, P., Ecological apes and ancestors, Nature 376:556, 1995. Natrag na tekst.
- Leakey, M.G., Feibel, C.S., McDougall, I., Ward, C. and Walker, A., New specimens and confirmation of an early age for Australopithecus anamensis, Nature 393:62, 1998; The distal end of the humerus was discovered in 1965 and is described in: Patterson, B. and Howells, W.W., Hominid humeral fragment from early Pleistocene of Northwest Kenya, Science 156:64–66, 1967. Natrag na tekst.
- Leakey et al., ref. 42, p. 62. Natrag na tekst.
- Richmond, B.G. and Strait, D.S., Evidence that humans evolved from a knuckle-walking ancestor, Nature 404:382, 2000. Natrag na tekst.
- Ward, C., Leakey, M. and Walker, A., The new hominid species Australopithecus anamensis, Evolutionary Anthropology 9:201, 1999. Natrag na tekst.
- Reader, J., Missing Links: The Hunt for Earliest Man, Penguin Books, London, pp. 207–210, 1988. Natrag na tekst.
- Lewin, R., Bones of Contention, Penguin Books, London, pp. 268–300, 1987. Natrag na tekst.
- Wood, B. and Richmond, B.G., Human evolution : taxonomy and paleobiology, J. Anatomy 196:29, 2000. Natrag na tekst.
- McHenry, H.M. and Coffing, K., Australopithecus to Homo: transformations in body and mind, Annual Review of Anthropology 29:127, 2000. Natrag na tekst.
- Tattersall, I. and Schwartz, J.H., Extinct Humans, Westview Press, New York, p. 88, 2001. Natrag na tekst.
- Wood and Richmond, ref. 48, p. 28. Natrag na tekst.
- Kimbel, W.H., Johanson, D.C. and Rak, Y., The first skull and other new discoveries of Australopithecus afarensis at Hadar, Ethiopia, Nature 368:449–451, 1994. Natrag na tekst.
- Aiello, L.C., Variable but singular, Nature 368:399, 1994. Natrag na tekst.
- Kimbel, W.H., White, T.D. and Johanson, D.C., Cranial morphology of Australopithecus afarensis: a comparative study based on a composite reconstruction of the adult skull, American J. Physical Anthropology 64:337–388, 1984; Kimbel, W.H. and White, T.D., A revised reconstruction of the adult skull of Australopithecus afarensis, J. Human Evolution 17:545–550, 1988. Natrag na tekst.
- Shreeve, J., ‘Lucy,’ crucial early human ancestor, finally gets a head, Science 264:34, 1994. Natrag na tekst.
- Leakey, R. and Lewin, R., Origins Reconsidered: In Search of What Makes us Human, Abacus, London, pp. 193–196, 1992. Natrag na tekst.
- Stern, J.T. and Susman, R.L., The locomotor anatomy of Australopithecus afarensis, American J. Physical Anthropology 60:284–285, 1983. Natrag na tekst.
- Cherfas, J., Trees have made man upright, New Scientist 97:172, 1983. Natrag na tekst.
- Richmond, B.G., Aiello, L.C. and Wood, B.A., Early hominin limb proportions, J. Human Evolution 43:534–535, 2002. Natrag na tekst.
- Tattersall and Schwartz, ref. 50, p. 90. Natrag na tekst.
- Richmond et al., ref. 59, pp. 542–543. Natrag na tekst.
- Stern and Susman, ref. 57, p. 284. Natrag na tekst.
- Stern and Susman, ref. 57, p. 307. Natrag na tekst.
- Stern and Susman, ref. 57, p. 308. Natrag na tekst.
- Stern and Susman, ref. 57, pp. 283–284. Natrag na tekst.
- Richmond and Strait, ref. 44, p. 383. Natrag na tekst.
- Richmond and Strait, ref. 44, pp. 383–384. Natrag na tekst.
- Cherfas, ref. 58, p. 176. Natrag na tekst.
- Collard, M. and Aiello, L.C., From forelimbs to two legs, Nature 404:339, 2000. Natrag na tekst.
- Gamble, C., Timewalkers: The Prehistory of Global Colonization, Penguin Books, London, p. 58, 1993. Natrag na tekst.
- Abitbol, M.M., Lateral view of Australopithecus afarensis: primitive aspects of bipedal positional behavior in the earliest hominids, J. Human Evolution 28:228, 1995. Natrag na tekst.
- Abitbol, ref. 71, p. 211. Natrag na tekst.
- Berge, C., How did australopithecines walk? A biomechanical study of the hip and thigh of Australopithecus afarensis, J. Human Evolution 26:270, 1994. Natrag na tekst.
- Berge, ref. 73, p. 271. Natrag na tekst.
- Stern and Susman, ref. 57, p. 313. Natrag na tekst.
- Shipman, P., Those ears were made for walking, New Scientist 143:27, 1994. Natrag na tekst.
- Spoor, F., Wood, B. and Zonneveld, F., Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion, Nature 369:645, 1994. Natrag na tekst.
- Gibbons, A., Anthropologists take the measure of humanity, Science 264:350, 1994. Natrag na tekst.
- Lubenow, M.L., Paleoanthropology in review, Journal of Creation 10(1):13, 1996. Natrag na tekst.
- Kohler, M. and Moya-Sola, S., Ape-like or hominid-like? The positional behavior of Oreopithecus bambolii reconsidered, Proc. Nat. Acad. Sci. USA 94:11747–11750, 1997. Natrag na tekst.
- Kohler and Moya-Sola, ref. 80, p. 11747. Natrag na tekst.
- Gee, H., Return to the planet of the apes, Nature 412:131, 2001. Natrag na tekst.
- Leakey, M.G., Spoor, F., Brown, F.H., Gathogo, P.N., Klarie, C., Leakey, L.N. and McDougall, I., New hominin genus from eastern Africa shows diverse middle Pliocene lineages, Nature 410:433–440, 2001. Natrag na tekst.
- Wong, K., Finding Homo sapiens’ lost relatives, Scientific American 285(4):26–27, 2001. Natrag na tekst.
- White, T., Early hominids—diversity or distortion? Science 299:1994–1997, 2003. Natrag na tekst.
- Brunet, M., Beauvilain, A., Coppens, Y., Heintz, E., Moutaye, A.H.E. and Pilbeam, D., The first australopithecine 2,500 kilometres west of the Rift Valley (Chad), Nature 378:273–275, 1995. Natrag na tekst.
- Falk, D., Redmond Jr, J.C., Guyer, J., Conroy, G.C., Recheis, W., Weber, G.W. and Seidler, H., Early hominid brain evolution: a new look at old endocasts, J. Human Evolution 38:709, 2000. Natrag na tekst.
- Klein, R.K. and Edgar, B., The Dawn of Human Culture, John Wiley & Sons Inc., New York, p. 35, 2002. Natrag na tekst.
- Wood and Richmond, ref. 48, p. 32. Natrag na tekst.
- McHenry, H.M. and Berger, L.R., Body proportions in Australopithecus afarensis and A. africanus and the origin of the genus Homo, J. Human Evolution 35:1–22, 1998; McHenry, H.M. and Berger, L.R., Limb lengths in Australopithecus and the origin of the genus Homo, South African J. Science, 94:447–450, 1998. Natrag na tekst.
- McHenry and Berger, ref. 90, p. 447. Natrag na tekst.
- Klein and Edgar, ref. 88, p. 37. Natrag na tekst.
- McHenry, H.M., The first bipeds: a comparison of the A. afarensis and A. africanus postcranium and implications for the evolution of bipedalism, J. Human Evolution 15:179, 1986. Natrag na tekst.
- McHenry, ref. 93, p. 186. Natrag na tekst.
- McHenry, ref. 93, p. 187. Natrag na tekst.
- Asfaw, B., White, T., Lovejoy, O., Latimer, B., Simpson, S. and Suwa, G., Australopithecus garhi: a new species of early hominid from Ethiopia, Science 284:631, 1999. Natrag na tekst.
- Asfaw et al., ref. 96, pp. 632–633. Natrag na tekst.
- Richmond et al., ref. 59, p. 542. Natrag na tekst.
- Richmond et al., ref. 59, p. 543. Natrag na tekst.
- McCollum, M.A., The robust australopithecine face: a morphogenetic perspective, Science 284:301, 1999. Natrag na tekst.
- Johanson, D. and Edgar, B., From Lucy to Language, Simon & Schuster Editions, New York, p.152, 1996. Natrag na tekst.
- Klein and Edgar, ref. 88, p. 41. Natrag na tekst.
- Klein and Edgar, ref. 88, p. 39. Natrag na tekst.
- Johanson and Edgar, ref. 101, pp. 23, 152. Natrag na tekst.
- Klein and Edgar, ref. 88, pp. 57–58. Natrag na tekst.
- Mehlert, A.W., A review of the present status of some alleged early hominds, Jornal of Creation 6(1):32, 1992. Natrag na tekst.
- Mehlert, ref. 106, p. 34. Natrag na tekst.
- Johanson and Edgar, ref. 101, p. 23. Natrag na tekst.
- Falk et al., ref. 87, pp. 708–709. Natrag na tekst.
- Wood and Richmond, ref. 48, p. 37. Natrag na tekst.
- Wood and Richmond, ref. 48, p. 38. Natrag na tekst.
- Wood and Richmond, ref. 48, p. 35. Natrag na tekst.
- Mehlert, A.W., Australopithecines—the extinct southern apes of Africa: a fresh light on their status? Journal of Creation 14(3):91–99, 2000.Natrag na tekst.
- Walker, A. and Shipman, P., The Wisdom of Bones: In Search of Human Origins, Weidenfeld and Nicolson, London, pp. 192, 1996. Natrag na tekst.



Readers’ comments
Comments are automatically closed 14 days after publication.